Potato brown rot was classified as a quarantine disease because of the complexity of the pathogen Ralstonia solanacearum L., formerly called Pseudonomas or Burkholderia solanacearum. This same bacterium causes the disease bacterial wilt. It is a multifaceted bacterium hence the complex name Ralstonia solanacearum (CeRs or RASLSSO) or BW (in English). It attacks several plant families, notably nightshades such as potatoes; the tomato; eggplant; chili pepper; tobacco…etc. The potato is the most cultivated plant worldwide. Mali is the second largest producer in West Africa after Nigeria. It imports more than 9,000 to 11,000 tons of seeds each year. Potato cultivation is faced with several biotic and abiotic attacks. Among all these attacks, those caused by bacteria cause the most significant damage. In addition, diseases caused mainly by the Ralstonia solanacearum complex lack adequate control solutions. Despite everything, today, very few studies are carried out to identify these bacteria in order to prevent their spread in the soil in Mali. This is why the objective of this study is to identify the bacteria responsible for brown rot in potato seeds in Mali. To achieve our objective, a total of 63 samples were taken from nine varieties of imported and locally multiplied potato seeds. The agent responsible for brown rot was isolated on the Triphenyl-Tetrazolium-chloride culture medium, and several microbiological, biochemical and molecular tests were carried out to identify the different isolates selected. As the main results of this study: twenty-seven bacterial isolates, capable of causing brown rot of potato seeds were isolated, the biochemical tests carried out on these isolates showed that they all of race3 and twenty -four isolates were biovar 3; one from biovar 2 and two from biovar 6. The molecular studies carried out using universal primers confirmed the presence of the species Ralstonia solanacearum with two phylotypes (I and II).
| Published in | International Journal of Microbiology and Biotechnology (Volume 9, Issue 3) |
| DOI | 10.11648/j.ijmb.20240903.11 |
| Page(s) | 43-53 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Seeds, Potato, Brown Rot, Ralstonia Solanacearum, PCR
Substrates | Biovars | ||||||
|---|---|---|---|---|---|---|---|
1 | 2 | 3 | 4 | 5 | 6 | 3b | |
Cellobiose | - | + | + | - | + | + | + |
Maltose | - | + | + | - | + | + | + |
Lactose | - | + | + | - | + | + | - |
Dulcitol | - | - | + | + | - | - | - |
Sorbitol | - | - | + | + | - | + | + |
Mannitol | - | - | + | + | + | + | + |
Primer names | Sequences 5'-3' | Amplified fragment size | Phylotypes | |
|---|---|---|---|---|
Nmult | 21:1F | CGTTGATGAGGCGCGCAATTT | 144 bp | I (Asiaticum) |
Nmult | 21:2F | AAGTTATGGACGGTGGAAGTC | 372 bp | II (Americanum) |
Nmult | 23:AF | ATTACSAGAGCAATCGAAAGATT | 91 bp | III (African) |
Nmult | 22:InF | ATTGCCAAGACGAGAGAAGTA | 213pb | IV (Tropical) |
Nmult | 22:RR | TCGCTTGACCCTATAACGAGTA | Reverse primer | Any phylotype |
759 | R | GTCGCCGTCAGCAATGCGGAATCG | 280 bp (Internal marker) | Ralstonia |
760 | F | GTCGCCGTCAGCAATGCGGAATCG | ||
Source of seeds | Potato host varieties | Name of isolates | Total number |
|---|---|---|---|
Imported | Claustar | RIC1 | 13 |
Mondial | RIM1, RIM2 | ||
Liseta | RIL1, RIL2 | ||
Sagitta | RISa1, RISa2 | ||
Sifra | RISi1 | ||
Spunta | RISp1 | ||
Evora | RIE1, RIE2 | ||
El Beida | RIEL1, RIEL2 | ||
Local | Sahel | RLSh1 | 18 |
Claustar | RLC1, RLC2, RC3, RLC4, RLC5, RLC6, RLC7, RLC8, RLC9, RLC10 | ||
Spunta | RLSp1, RLSp2, RLSp3, RLSp4 RLSp5, RLSp6 RLSp7 | ||
Total | 31 |
Isolates tested | Testing | ||||
|---|---|---|---|---|---|
KOH | Gram | Catalase | Oxidase | Fluorescent pigment production | |
RISa1; RIE1; RIM1; RIEL1; RLSh1; RLSp1; RLSp2; RLSp3; RLSp4; RLC3; RLC6; RLC7; RLC8; RLC9; RLC10; RLSp7 | + | - | + | + | - |
RISi1; RISp1; RIE2; RISa2; RIEL2; RLC1; RLC2; RLC4; RLC5; RLSp5; RLSp6 | + | - | + | - | - |
Isolates tested | Number of plants infiltrated per isolate | Number of wilted plants per infiltrated isolate |
|---|---|---|
RISa1; RISi1; RISa2; RISp1; RIE1; RIEL1; RIEL2; RLSh1, RLC1; RLC2; RLC4; RLC5; RLC6; RLC7; RLC8; RLC10; RLSp1; RLSp2; RLSp3; RLSp4; RLSp5; RLSp7 | 3 | 3 |
RIE2; RIM1; RLC3; RLC9; RLSp6 | 3 | 0 |
Isolates tested | Disaccharides | Hexose alcohols | Biovars | ||||
|---|---|---|---|---|---|---|---|
Cellobiose | Lactose | Maltose | Mannitol | Dulcitol | Sorbitol | ||
RLC6 | + | + | + | - | - | - | 2 |
RISa1; RISi1; RISp1; RIE1; RIE2; RISa2; RIEL1; RIEL2; RLSh1; RLSp1; RLSp2; RLC1; RLSp3; RLSp4; RLC2; RLC3; RLC4; RLC5; RLC7; RLSp5; RLSp6; RLC8; RLC9; RLSp7 | + | + | + | + | + | + | 3 |
RIM1; RLC10 | + | + | + | + | - | + | 6 |
Isolates | Phylotypes |
|---|---|
RLSp6; RLC9 | I |
RISa1; RISi1; RISp1; RIE1; RIE2; RIM1; RISa2; RIEL1; RIEL2; RLSh1; RLSp1; RLSp2; RLC1; RLSp3; RLSp4; RLC2; RLC3; RLC4; RLC5; RLC6; RLC7; RLSp5; RLC8; RLC10; RLSp7 | II |
PCR | Polymerization Chain Reaction |
UV | Ultraviolet |
Pb | Pair of Bands |
Labosem-Sotuba | Plant Seed Certification Laboratory |
| [1] | COAG/2022/19. Proposal relating to the proclamation of an International Potato Day. |
| [2] | FaoStat. (2018). Prospective analysis bulletin of the agricultural market. SOS potato sector. Crops and animal products. 12 p. |
| [3] | Inter-professional Potato Group. 2019. newsletter N°2, rap pdt, 2017; bio online, 2019. |
| [4] | Bruno Vanderhofstad (2019). Information and status report on the production of seed potatoes in Mali. Return of information to the Sikasso workshop. |
| [5] | Potato Mali Interprofessional Association. (2021). National strategy for the development of the potato sector in Mali. Produce 1 million tonnes by 2026. Article by Brehima Diallo. |
| [6] | Moussa BOIRE and Lassana B. TRAORE. (2018). Identification of Erwinia responsible for soft rot of potatoes sold on the Medina Coura market. (Bamako). 1. P. 9. |
| [7] | Rakotoniaina Andry Mampionona. (2022). Exploitation of soil Actinomycetes for the biocontrol of potato bacteriosis caused by Ralstonia solanacearum. 83 p. |
| [8] | FaoStat. (2020). Report of the Committee on Agriculture, twenty-eighth sessions. Proposal relating to the proclamation of an international potato day. |
| [9] | Florent AILLOUD. (2015). Pathogenicity in Ralstonia solanacearum phylotype II integrative genomics and transcriptomic landscapes in relation to host adaptation. Thesis: UNIVERSITY OF RÉUNION Faculty of Sciences and Technologies Doctoral School of Sciences, Technologies and Health. 11. P (223). |
| [10] | Salgon, S. (2017). Genetic determinism of resistance to bacterial wilt in eggplant and applications in varietal selection (Doctoral dissertation, University of Reunion). |
| [11] | Kelman A. 1954. The relationship of pathogenicity in Pseudomonas solanacearum to colony appearance on a tetrazolium medium. Phytopathology 44: 693–695. |
| [12] | Kassogué et al (2015). Isolation and characterization of Bacillus thuringiensis (Ernst Berliner) strains indigenous to agricultural soils of Mali, pages n°6. 9 p. |
| [13] | Zubeda, C. and Hamid, R. (2011). Isolation and characterization of Ralstonia solanacearum from infected tomato plants of Soan Skesar valley of Punjab. Pak. J. Bot. 7(2). P. 2980. |
| [14] | RR Bhanwar, PK Tiwari, RK Dantre, RR Saxena, A. Tiwari and A. Kotasthane. (2021). Determination of Races and Biovars of Ralstonia solanacearum causing Bacterial Wilt of Brinjal in Chhattisgarh, India, International Journal of Current Microbiology and Applied Sciences. ISSN: 2319-7706 Volume 10 Number 06. (2021). 5-6. 12P. |
| [15] | Hayward, A. C., (1964). Characteristics of Pseudomonas solanacearum. J Appl Microbiol. 27(2): 265-277. |
| [16] | HaywardAC, El-Nashaar HM, Nydegger U. and DE Lindo L. 1990. Variation in nitrate metabolism in biovars of Pseudomonas solanacearum. Journal of Applied Bacteriology, Vol. 69, p. 269-280. |
| [17] | Hayward AC, 1976. Systematics and relationship of Pseudomonas solanacearum, p. 6-21. In: Proceedings of the 1st International Conference and Workshop on the Ecology and Control of Bacterial Wilt Caused by Pseudomonas solanacearum. L. Sequeira and A. Kelman (Editors), North Carolina State University, Raleigh, NC. |
| [18] | Martin, Prior, P., Cadet, P., & Steva, H. (1989). Variability of the pathogenic power of Pseudomonas solanacearum in the French West Indies (Martinique and Guadeloupe). Acta Oecol, 10, 135-142. |
| [19] | Schaad NW, Jones JB, Chun W. (2001). Laboratory guide for the identification of plant pathogenic bacteria. 3rd ed. APS Press, St. Paul, USA. p 373. |
| [20] |
PLOS UN
https://doi.org/10.1371/journal.pone.0274780 September 22, 2022 3 / 36. |
| [21] | Lozano, J. and Sequeira, L. (1970). Differentiation of Ralstonia solanacearum races using a foliar infiltration technique. Phytopathol. 60. P. 833-838. |
| [22] | Fegan and Prior. (2005). Ralstonia solanacearum species complex”. In Allen Bacterial wilt disease and the Ralstonia solanacearum species. |
| [23] | Opina N., F. Tavner, G. Hollway, J.-F. Wang, T.-H. Li, R. Maghirang, M. Fegan, AC Hayward, V. Krishnapillai, WF Hong, BW Holloway, and J. Timmis. (1997). A novel method for development of species and strain-specific DNA probes and PCR primers for identifying Burkholderia solanacearum (formerly Pseudomonas solanacearum). Asia Pac. J. Mol. Biol. Biotechnology. 5: 19–30. |
| [24] | Ravelomanantsoa Santatra Herilalaina, (2016). Population biology of the Ralstonia solanacearum species complex applied to the epidemiology of bacterial wilt of potatoes in Madagascar. PhD. University of Antananarivo. Doctoral School of Life and Environmental Sciences. Biotechnology specialty. Antananarivo. Madagascar. 27-28. P (239). |
| [25] | Adamou (2011), Characterization of local strains of Ralstonia solanacearum (EF SMITH) YABUUCHI et al. and evaluation of the behavior of six varieties of potato (Solanum tuberosum L.) against strain RS-09-76, in Niger. Doctoral thesis. University of Ouagadougou. P (76-89), 197P. |
| [26] | Aïssa Thera (2007), Bacterial wilt management: a prerequisite for a potato seed certification program in Mali, Master of Science Thesis. Montana State University. USA. P (94-97). 142P. |
| [27] | Rakotoniaina Andry M. (2022). Epidemiological study of bacterial wilt of potatoes in the Vakinankaratra region of Madagascar. P (26-30). 83P. |
| [28] | Parrur. (2013). Integrated control of bacterial wiltof bacteriosis (Ralstonia solanacearum) of potatoes in small farms. through the control of the phytosanitary state of potato seed tubers and the detection of populations of Ralstonia solanacearum in the soil of the Vakinankaratra Region. 13-15. P (40). |
| [29] | Magnan Diarraand hasl (2019). Search for microorganisms capable of controlling strains of Ralstonia solanacearumisolated and characterized in the Baguinéda Irrigated Perimeter Office. article of. |
| [30] | Ankit Kumar Ghorai, Subrata Dutta. Ashis Roy BarmanID (2022). Genetic diversity of Ralstonia solanacearum causing vascular bacterial wilt under different agro-climatic regions of West Bengal, India, 8-9; 36p. |
| [31] | Hayward, A. (1991). Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Anne. everend Phytopathol. 29. P. 65-87. |
| [32] | Leroux AC. (1996). Study of the resistance of potato varieties and genotypes to Ralstonia solanacearum, In: Production of serological reagents for the detection of Ralstonia solanacearum. INRA-FNPPPT activity report. p. 27-31. |
| [33] | Wilfried Kayaba Ouédraogo. (2016). Characterization of isolates of Ralstonia solanacearum (EFSMITH), Yabuuchi et al, causal agent of bacterial wilt in the market gardening areas of Houet, Kenedougou and Passore and Search for varieties resistant to the disease in Burkina Faso, End of cycle dissertation, Institute Polytechnique de Bobo-Dioulasso, 9-10, P. 66. |
| [34] | Popoola, Arizona, Ganiyu, SA, Enikuomehin, OA, Bodunde, JG, Adedibu, O B., Durosomo, HA and Karunwi, OA. (2015). Isolation and characterization of Ralstonia solanacearum Causing bacterial wilt of tomato in Nigeria. 6-7. P. 10. |
| [35] | Castillo, JA and Greenberg, JT (2007). Evolutionary dynamics of Ralstonia solanacearum. Appl. Approximately. Microbiol. 73. 1225–1238. |
| [36] |
Wicker E, Lefeuvre P, de Cambiaire JC, Lemaire C, Poussier S, Prior P. (2012). Contrasting recombination patterns and demographic histories of the plant pathogen Ralstonia solanacearum inferred from MLSA. ISME Magazine. 6(5): 961-974 from
https://doi.org/10.1038/ismej.2011.160 PM ID: 22094345. |
| [37] | Hasina Ny Aina Rasoamanana. (2022). Population biology of the Ralstonia solanacearum species complex in Madagascar and the southwest Indian Ocean. Thesis. Hal. Science. 45(32). P (242). |
APA Style
Diallo, B., Kassogué, A., Fané, R., Coulibaly, M., Dao, S., et al. (2024). Diversity of the Ralstonia Solanacearum Species of Potato Seeds in Mali. International Journal of Microbiology and Biotechnology, 9(3), 43-53. https://doi.org/10.11648/j.ijmb.20240903.11
ACS Style
Diallo, B.; Kassogué, A.; Fané, R.; Coulibaly, M.; Dao, S., et al. Diversity of the Ralstonia Solanacearum Species of Potato Seeds in Mali. Int. J. Microbiol. Biotechnol. 2024, 9(3), 43-53. doi: 10.11648/j.ijmb.20240903.11
AMA Style
Diallo B, Kassogué A, Fané R, Coulibaly M, Dao S, et al. Diversity of the Ralstonia Solanacearum Species of Potato Seeds in Mali. Int J Microbiol Biotechnol. 2024;9(3):43-53. doi: 10.11648/j.ijmb.20240903.11
@article{10.11648/j.ijmb.20240903.11,
author = {Binta Diallo and Adounigna Kassogué and Rokiatou Fané and Moctar Coulibaly and Sognan Dao and Ibrahima Mallé and Amadou Hamadoun Babana},
title = {Diversity of the Ralstonia Solanacearum Species of Potato Seeds in Mali
},
journal = {International Journal of Microbiology and Biotechnology},
volume = {9},
number = {3},
pages = {43-53},
doi = {10.11648/j.ijmb.20240903.11},
url = {https://doi.org/10.11648/j.ijmb.20240903.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijmb.20240903.11},
abstract = {Potato brown rot was classified as a quarantine disease because of the complexity of the pathogen Ralstonia solanacearum L., formerly called Pseudonomas or Burkholderia solanacearum. This same bacterium causes the disease bacterial wilt. It is a multifaceted bacterium hence the complex name Ralstonia solanacearum (CeRs or RASLSSO) or BW (in English). It attacks several plant families, notably nightshades such as potatoes; the tomato; eggplant; chili pepper; tobacco…etc. The potato is the most cultivated plant worldwide. Mali is the second largest producer in West Africa after Nigeria. It imports more than 9,000 to 11,000 tons of seeds each year. Potato cultivation is faced with several biotic and abiotic attacks. Among all these attacks, those caused by bacteria cause the most significant damage. In addition, diseases caused mainly by the Ralstonia solanacearum complex lack adequate control solutions. Despite everything, today, very few studies are carried out to identify these bacteria in order to prevent their spread in the soil in Mali. This is why the objective of this study is to identify the bacteria responsible for brown rot in potato seeds in Mali. To achieve our objective, a total of 63 samples were taken from nine varieties of imported and locally multiplied potato seeds. The agent responsible for brown rot was isolated on the Triphenyl-Tetrazolium-chloride culture medium, and several microbiological, biochemical and molecular tests were carried out to identify the different isolates selected. As the main results of this study: twenty-seven bacterial isolates, capable of causing brown rot of potato seeds were isolated, the biochemical tests carried out on these isolates showed that they all of race3 and twenty -four isolates were biovar 3; one from biovar 2 and two from biovar 6. The molecular studies carried out using universal primers confirmed the presence of the species Ralstonia solanacearum with two phylotypes (I and II).
},
year = {2024}
}
TY - JOUR T1 - Diversity of the Ralstonia Solanacearum Species of Potato Seeds in Mali AU - Binta Diallo AU - Adounigna Kassogué AU - Rokiatou Fané AU - Moctar Coulibaly AU - Sognan Dao AU - Ibrahima Mallé AU - Amadou Hamadoun Babana Y1 - 2024/07/08 PY - 2024 N1 - https://doi.org/10.11648/j.ijmb.20240903.11 DO - 10.11648/j.ijmb.20240903.11 T2 - International Journal of Microbiology and Biotechnology JF - International Journal of Microbiology and Biotechnology JO - International Journal of Microbiology and Biotechnology SP - 43 EP - 53 PB - Science Publishing Group SN - 2578-9686 UR - https://doi.org/10.11648/j.ijmb.20240903.11 AB - Potato brown rot was classified as a quarantine disease because of the complexity of the pathogen Ralstonia solanacearum L., formerly called Pseudonomas or Burkholderia solanacearum. This same bacterium causes the disease bacterial wilt. It is a multifaceted bacterium hence the complex name Ralstonia solanacearum (CeRs or RASLSSO) or BW (in English). It attacks several plant families, notably nightshades such as potatoes; the tomato; eggplant; chili pepper; tobacco…etc. The potato is the most cultivated plant worldwide. Mali is the second largest producer in West Africa after Nigeria. It imports more than 9,000 to 11,000 tons of seeds each year. Potato cultivation is faced with several biotic and abiotic attacks. Among all these attacks, those caused by bacteria cause the most significant damage. In addition, diseases caused mainly by the Ralstonia solanacearum complex lack adequate control solutions. Despite everything, today, very few studies are carried out to identify these bacteria in order to prevent their spread in the soil in Mali. This is why the objective of this study is to identify the bacteria responsible for brown rot in potato seeds in Mali. To achieve our objective, a total of 63 samples were taken from nine varieties of imported and locally multiplied potato seeds. The agent responsible for brown rot was isolated on the Triphenyl-Tetrazolium-chloride culture medium, and several microbiological, biochemical and molecular tests were carried out to identify the different isolates selected. As the main results of this study: twenty-seven bacterial isolates, capable of causing brown rot of potato seeds were isolated, the biochemical tests carried out on these isolates showed that they all of race3 and twenty -four isolates were biovar 3; one from biovar 2 and two from biovar 6. The molecular studies carried out using universal primers confirmed the presence of the species Ralstonia solanacearum with two phylotypes (I and II). VL - 9 IS - 3 ER -
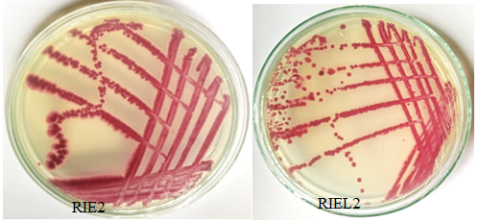
Figure 1. Macroscopic appearance of some isolates on TZC solid media.

Figure 2. Potato plants of the Naïma variety after inoculation with bacterial isolates. A: Overview of wilted plants 15 days after planting without infiltration, LaboSem-Sotuba, 2020; B: Wilting of plants of the Naïma variety 9 days after inoculation with the bacterial isolate RIEL2, LaboSem-Sotuba, 2020; C: Wilting of plants of the Naïma variety 22 days after inoculation with the bacterial isolate RLSp7 LaboSem-Sotuba, 2020.
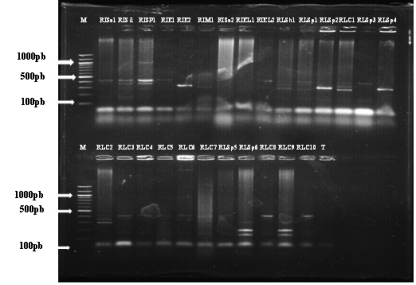
Figure 3. Migration profiles of Ralstonia solanacearum isolates amplified with the multiplex primers Nmult:21:1F (phylotype I), Nmult:21:2F (phylotype II), Nmult:23:AF (phylotype III), Nmult:22: InF (phylotype IV); Nmult:22:RR on AgaroseD1LOW EEO at 1.5%, migrated to 90V for 1 hour. After migration, the gel was photographed under UV using the Gel Doc device. RLSp6, RLC9 (phylotype I); RISa1; RISi1; RISp1; RIE1; RIE2;; RISa2; RIEL1; RIEL2; RLSp1; RLSp2; RLC1; RLSp3; RLSp4; RLC2; RLC3; RLC4; RLC5; RLC7; RLSp5; RLC8; RLSp7; RLC6; RIM1; RLC10 (phylotype II).
Information

